Continuity Nodes as Cybernetic Loops Evolutionary Rewiring, KEGGO Logic, and the Metabolic Cartography of Biological Nodes
Abstract
A conventional network node is a point. A Continuity Node is a point plus a loop: a site at which a system senses state, compares trajectories, modulates flux, and receives the consequences of its own modulation back into later behavior. Under this reading, the real unit of adaptive persistence is not the isolated node but the cybernetic cycle anchored there. This distinction matters because evolution does not merely favor connectivity. It favors architectures that can change without ceasing to function. Networks rewire because some loops absorb perturbation, some delay it, some redirect it, and some translate local shocks into system-wide correction. Continuity Nodes are thus best understood as the invariants of survivable change. Biological metabolism offers a privileged map of this principle. KEGG pathway maps can be read not only as reaction atlases but as landscapes of continuity-bearing junctions. Certain metabolites and regulatory axes matter so much because they preserve energy balance, redox buffering, branch-point allocation, chromatin state, and cell-fate option space while the broader network is being rearranged [1][2][3][4][5].
Full Text
SIGNALSENSE MONOGRAPH · CONTINUITY NODES DOSSIER
Continuity Nodes as Cybernetic Loops Evolutionary Rewiring, KEGGO Logic, and the Metabolic Cartography of Biological Nodes
A node matters not merely because many lines pass through it, but because a loop anchored there can keep the system coherent while the rest of the network changes.
This monograph reframes Continuity Nodes as loop-based control sites rather than static hubs. It argues that evolution favors such loops because they preserve viable passage through perturbation, allowing networks to rewire without systemic collapse. Read through KEGGO principles, biological metabolism appears as a map of continuity-bearing junctions: ATP/AMPK for energy continuity, NADPH/PPP for redox continuity, pyruvate for traffic switching, acetyl-CoA/citrate for carbon-to-identity coupling, and α-ketoglutarate–succinate–fumarate for epigenetic and cell-fate modulation. KEGG, in this view, is not only a pathway atlas but a cartography of cybernetic survivability.
SignalSense · Continuity Nodes Monograph · 1
Executive thesis
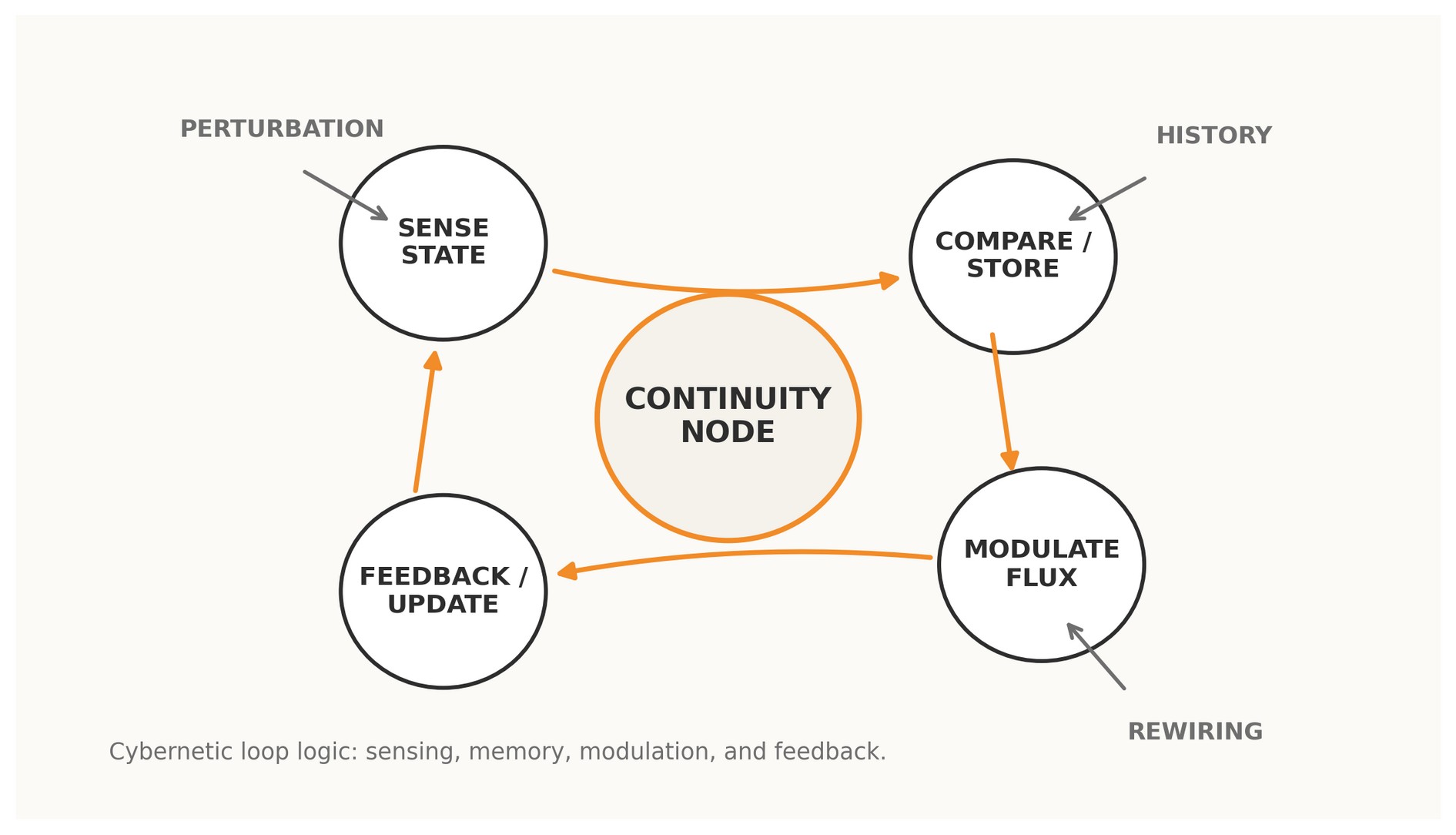
A conventional network node is a point. A Continuity Node is a point plus a loop: a site at which a system senses state, compares trajectories, modulates flux, and receives the consequences of its own modulation back into later behavior. Under this reading, the real unit of adaptive persistence is not the isolated node but the cybernetic cycle anchored there.
This distinction matters because evolution does not merely favor connectivity. It favors architectures that can change without ceasing to function. Networks rewire because some loops absorb perturbation, some delay it, some redirect it, and some translate local shocks into system-wide correction. Continuity Nodes are thus best understood as the invariants of survivable change.
Biological metabolism offers a privileged map of this principle. KEGG pathway maps can be read not only as reaction atlases but as landscapes of continuity-bearing junctions. Certain metabolites and regulatory axes matter so much because they preserve energy balance, redox buffering, branch-point allocation, chromatin state, and cell-fate option space while the broader network is being rearranged [1][2][3][4][5].
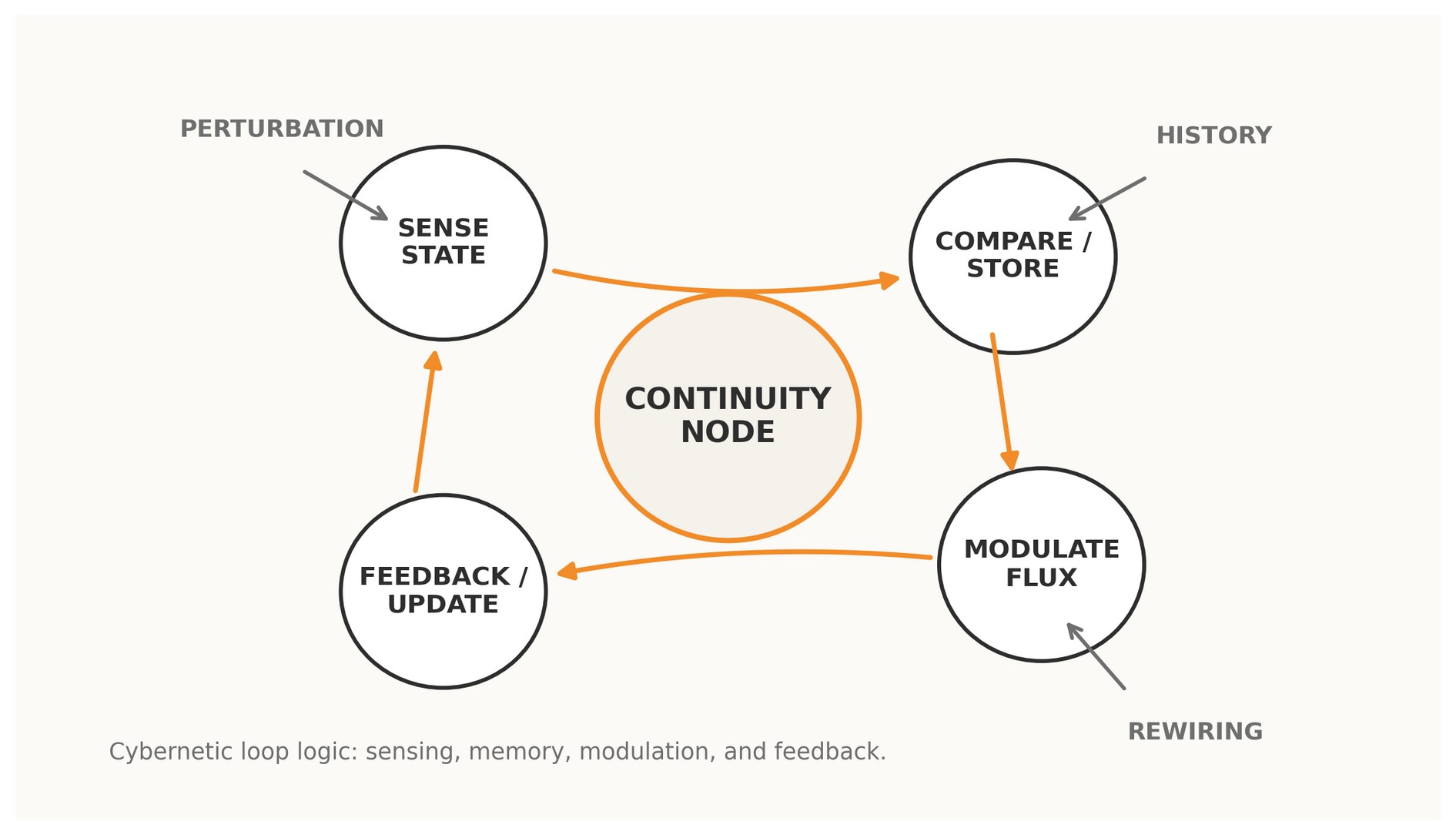
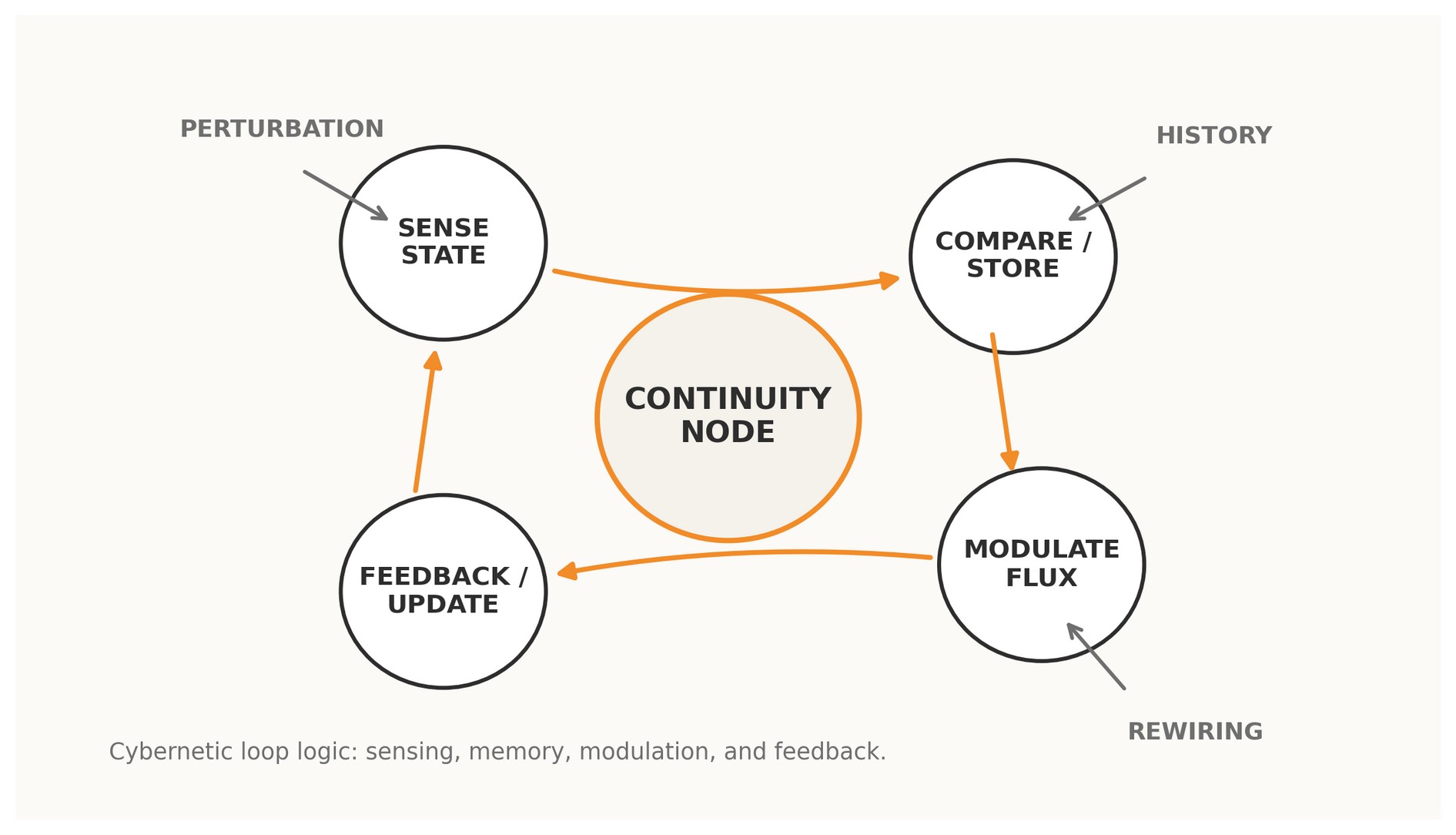
Figure 1. A Continuity Node read as a cybernetic loop: sensing, memory, modulation, feedback, and rewiring.
1 · Reframing nodes as loops
In graph-theoretic language, a node often appears as a static locus of adjacency. In a cybernetic language, that description is incomplete. A functioning node in a living or adaptive system is not defined only by how many edges touch it; it is defined by whether circular causality is organized around it. If the state sampled at the node changes how the network behaves, and if the consequences of that behavior return to shape subsequent states, the node is looped. If not, it is merely traversed.
The advantage of this reframing is explanatory. It distinguishes distribution from regulation. A hub may distribute traffic efficiently and still fail to stabilize a system under perturbation. By contrast, a continuity- bearing node can be modest in degree yet decisive for viability because it participates in a negative or layered feedback structure that keeps the system coherent under stress. Experimental work in synthetic biomolecular networks shows that layered feedback can outperform simpler controllers in the classic robustness–speed trade-off, demonstrating how multiple feedback strata can preserve response quality under disturbance [2].
SignalSense · Continuity Nodes Monograph · 2
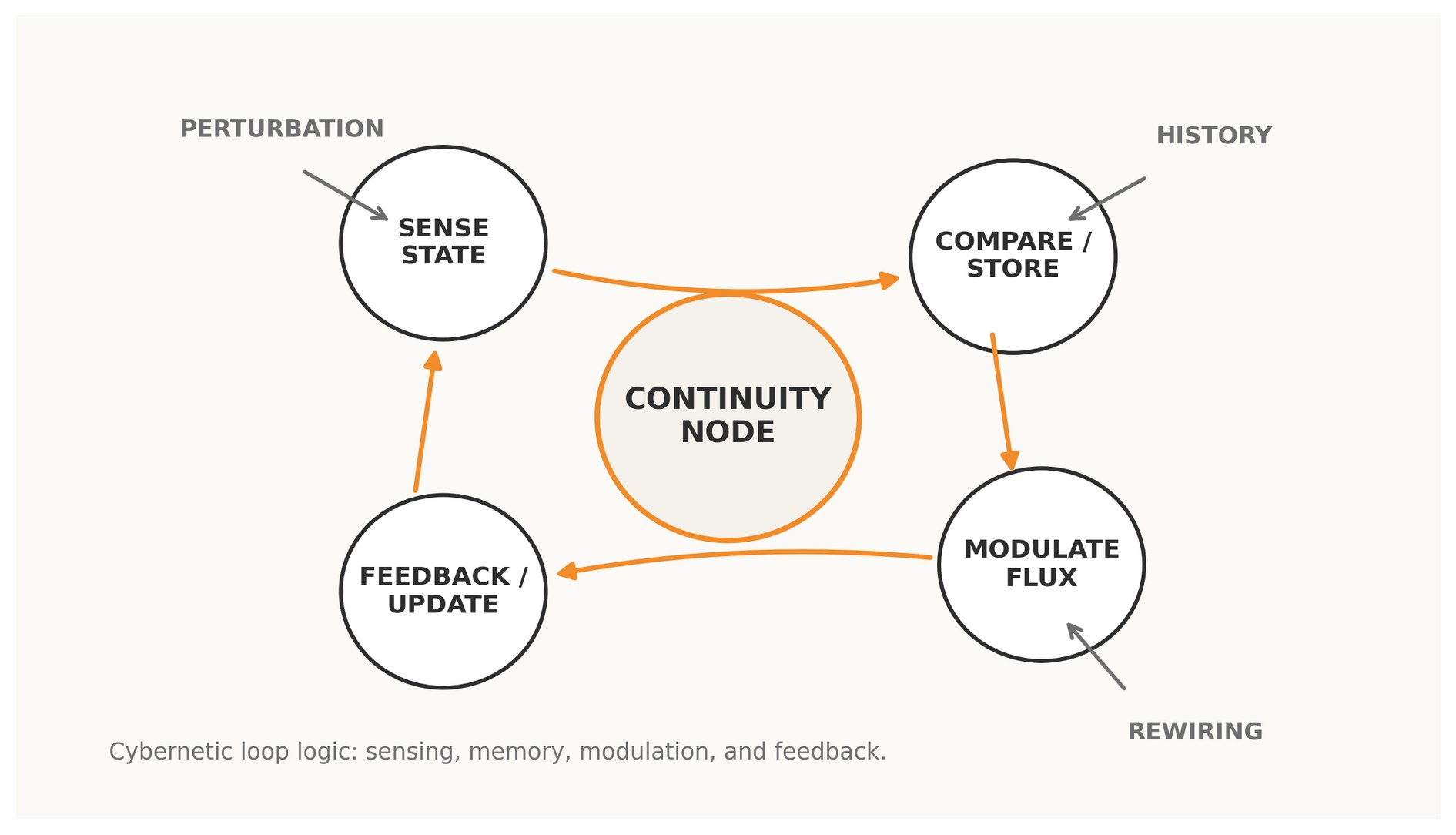
2 · Why loops govern evolutionary rewiring
Evolutionary rewiring is almost never an all-at-once replacement of topology. It is a sequence of local alterations—changes in interaction strengths, regulatory thresholds, expression patterns, branch-point preferences, or interspecies exchanges—that accumulate over time. Such accumulation is only possible if some parts of the network preserve function while others explore alternatives. Continuity Nodes are the structures that provide this buffering.
This is why rewiring and continuity are not opposites. They are reciprocals. A system can rewire because continuity loops absorb the first-order shock of mutation or environmental change; and continuity loops are retained because they allow the network to keep operating while its micro-architecture evolves.
Review work on biomolecular networks has emphasized that different biological networks are shaped by distinct selection pressures, and that their evolutionary behavior only becomes fully intelligible when topology is read together with temporal and spatial dynamics [6]. Adaptive laboratory evolution studies push the point further: growth restoration in genome-reduced Escherichia coli proceeded through broad transcriptome- and translatome-wide remodeling, coordinated by global regulatory changes rather than by a single isolated rescue mutation [3]. In other words, the rewiring that matters is often regulatory and systemic.
3 · KEGG as a cartography of continuity
KEGG PATHWAY is officially described as a collection of manually drawn pathway maps representing molecular interaction, reaction, and relation networks; its reference maps and derived organism-specific maps provide a standardized navigational substrate for metabolism, signaling, cellular processes, and disease [1]. That description is already powerful, but Continuity Nodes and KEGGO suggest a further reading.
A KEGG map can be treated not simply as a list of reactions but as a field of possible control junctions. The important positions are not always the most visually crowded ones. Rather, they are the sites at which flux allocation, sensing, redirection, and state preservation meet. A pathway atlas thus becomes a continuity atlas: a cartography of where the system can bend without breaking.
This reading is especially useful for metabolism because metabolic networks mix structure with thermodynamics. They are not only topological graphs; they are constrained flows of energy, matter, and signaling. Flux analysis has therefore become central to identifying regulatory leverage points, because it reveals which junctions actually redirect living physiology rather than merely appearing connected on paper [4].
SignalSense · Continuity Nodes Monograph · 3
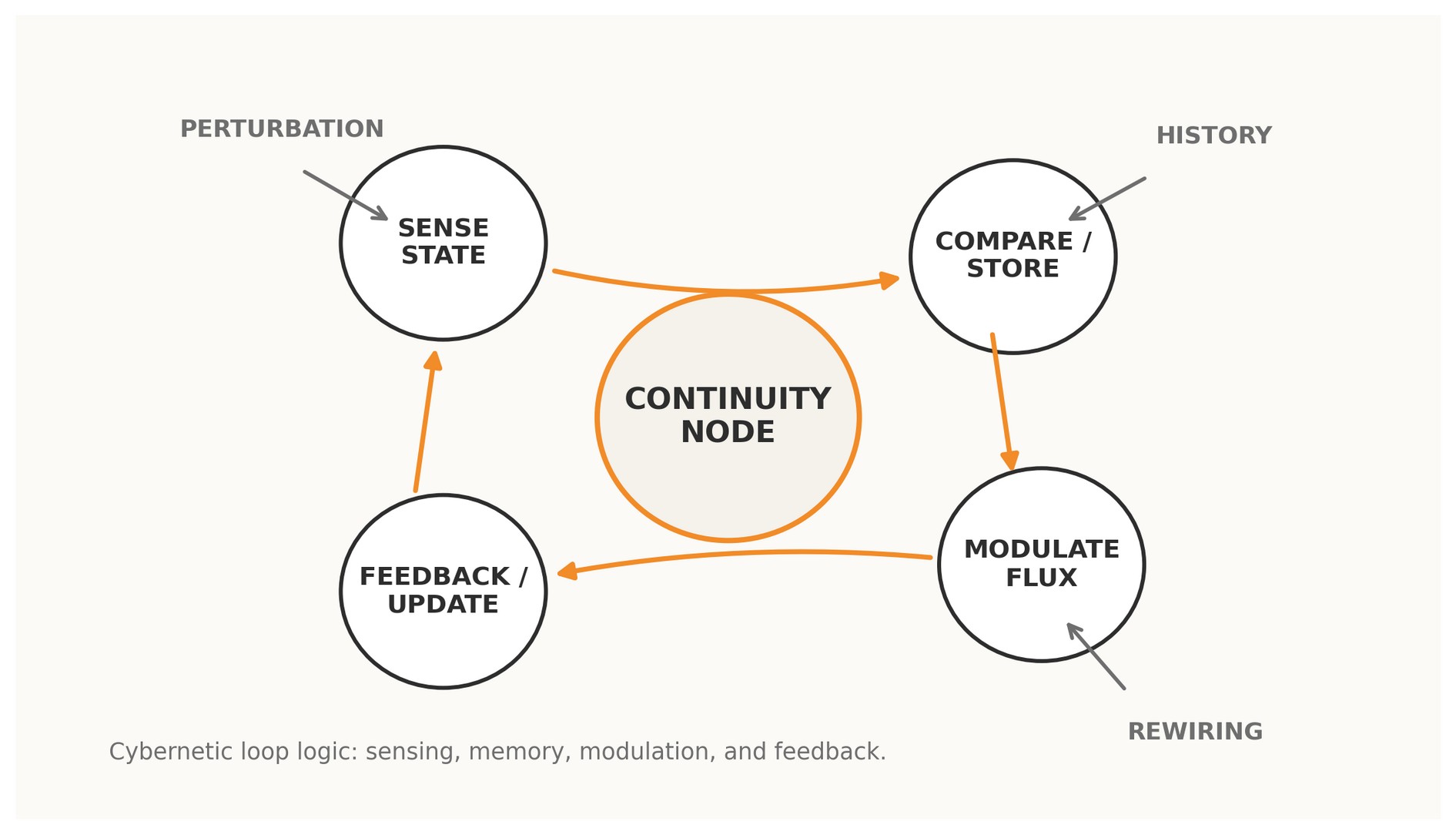
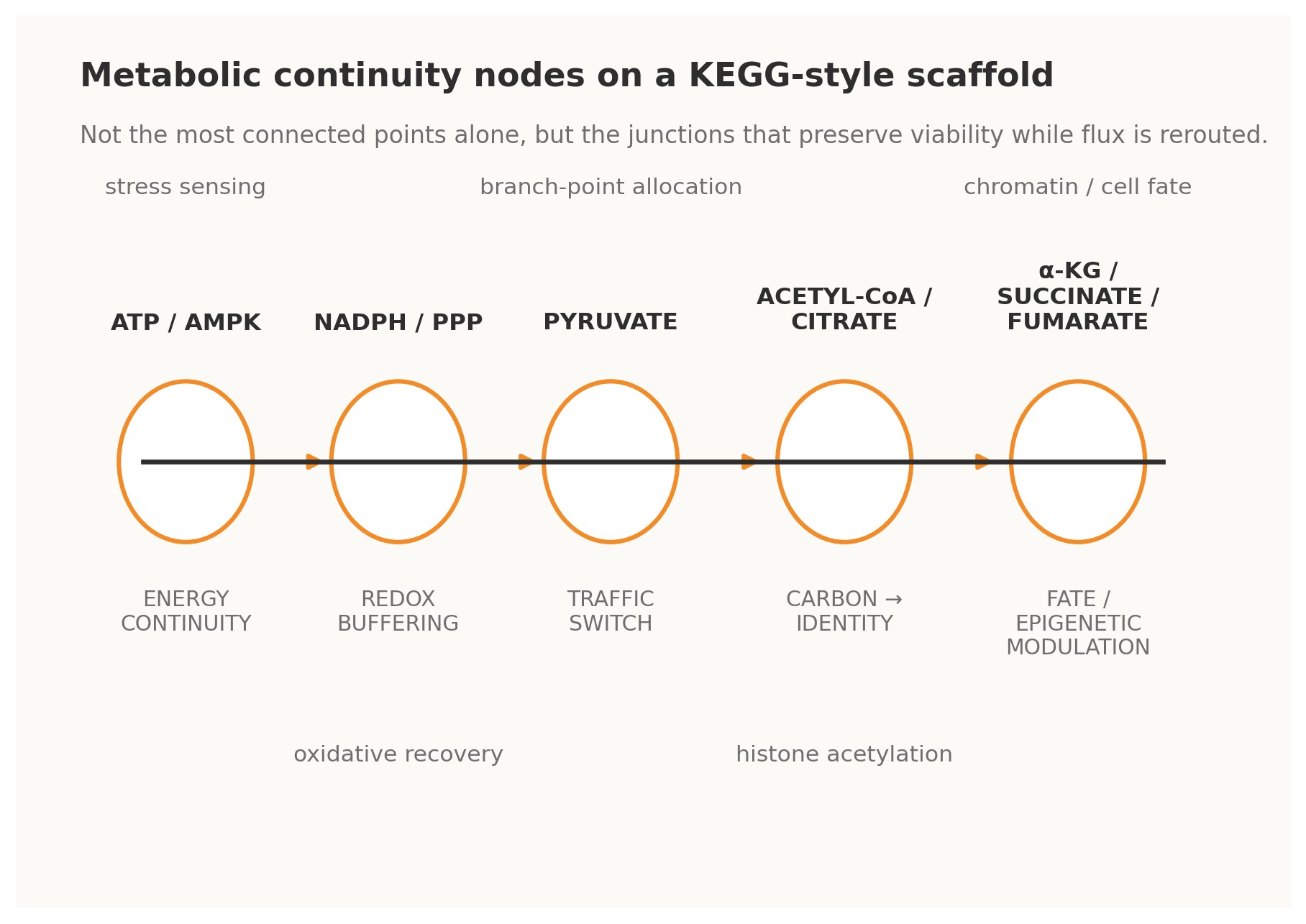
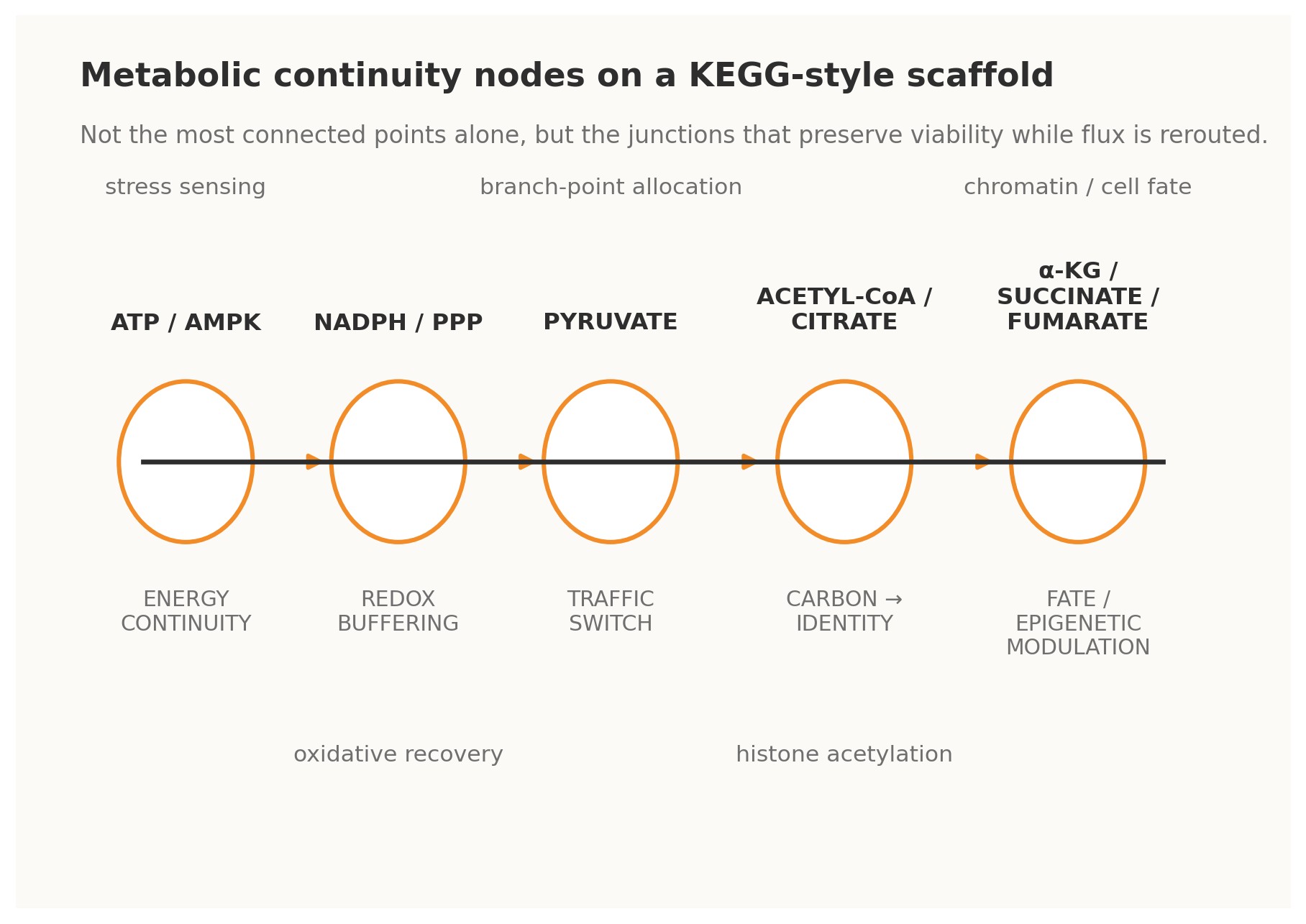
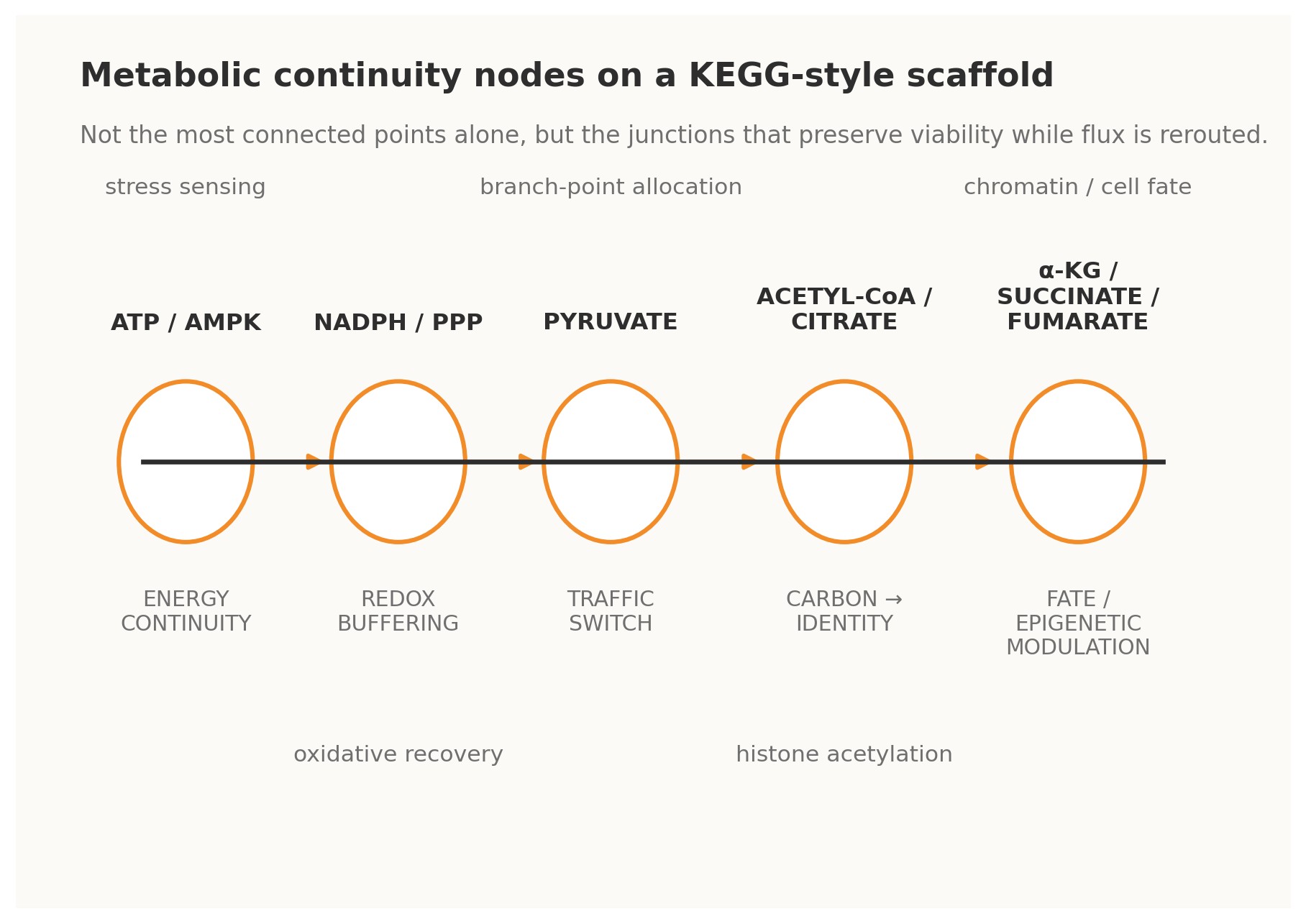
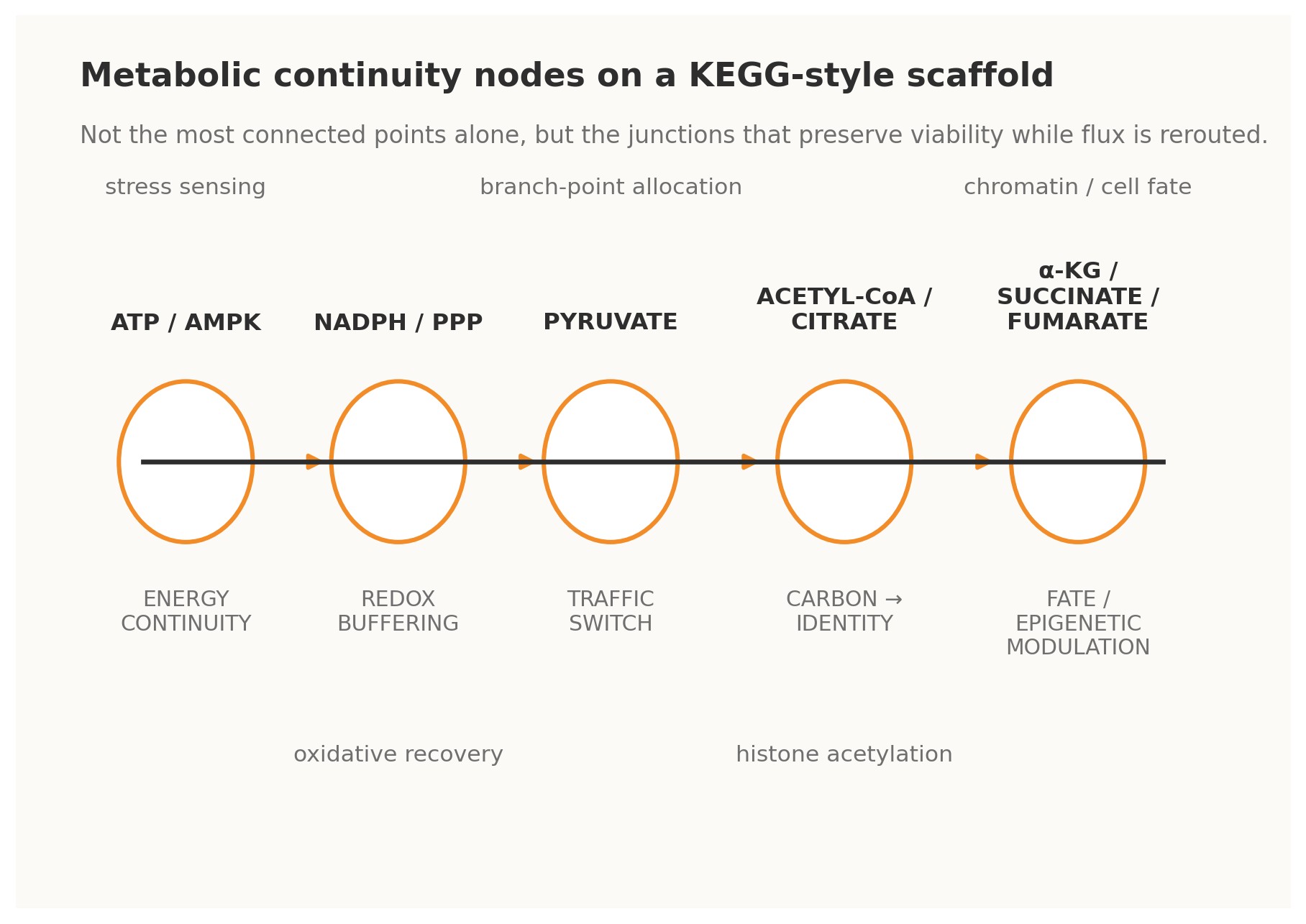
Figure 2. Metabolic analogues of Continuity Nodes on a KEGG-style scaffold.
4 · Canonical metabolic continuity nodes
4.1 · ATP / AMPK — energy continuity The ATP–ADP–AMP axis is perhaps the clearest metabolic example of a continuity-bearing loop. Changes in adenylate state do not merely report energetic strain; through AMPK and associated control systems they trigger compensatory shifts in uptake, biosynthesis, autophagy, and longer-horizon transcriptional programs. Energy status is therefore fed back into the operating logic of the cell. Under a Continuity Nodes reading, ATP/AMPK is not just a meter. It is a master governor that allows many other pathways to vary without immediate energetic collapse [4].
4.2 · NADPH / pentose phosphate pathway — redox continuity The oxidative pentose phosphate pathway behaves as a redox continuity module. When oxidative pressure rises and NADPH buffering becomes strained, flux can be redirected so that reducing power is regenerated. This is a classical cybernetic form: perturbation is sensed indirectly through redox state, the network reroutes carbon, and the resulting restored buffer capacity closes the loop. Such nodes matter evolutionarily because they enlarge the safe operating envelope for stress and mutation [4].
4.3 · Pyruvate — traffic-switch continuity Pyruvate sits at a decisive interface among respiration, fermentation, amino-acid exchange, and broader carbon allocation. Its importance lies not only in centrality but in optionality. It is a branch-point that can send metabolic commitment in multiple directions depending on need, oxygen availability, biosynthetic demand, or organismal context. In KEGGO language, pyruvate is a traffic-switch node: a gate through which distinct futures become differentially accessible [4].
4.4 · Acetyl-CoA and citrate — carbon-to-identity continuity Acetyl-CoA and citrate reveal that metabolism does not merely sustain the cell; it informs what the cell can become. Acetyl-CoA availability links carbon state to histone acetylation and broader chromatin regulation, while citrate export helps bridge mitochondrial metabolism to nuclear and cytosolic programs.
SignalSense · Continuity Nodes Monograph · 4
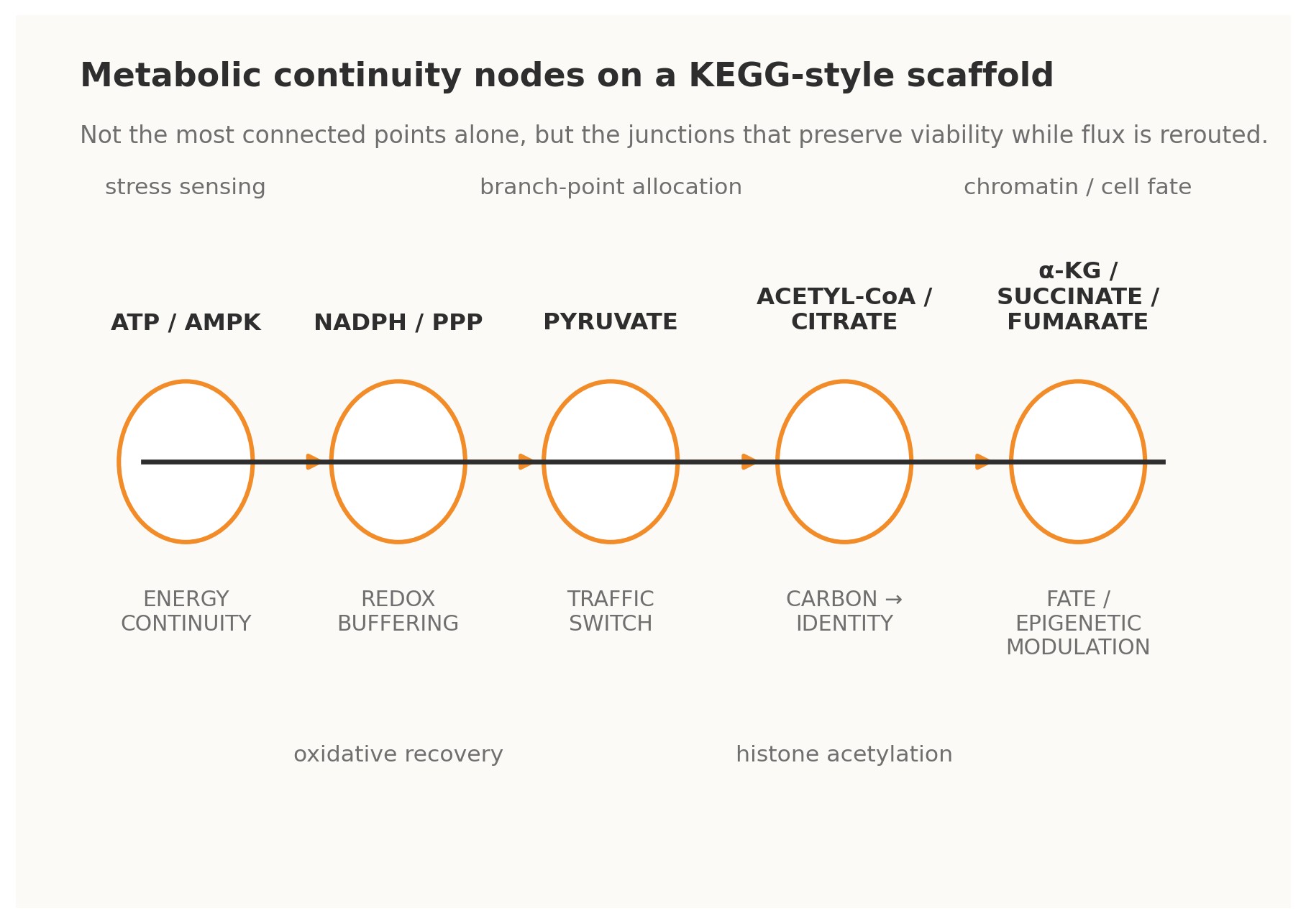
These nodes thus connect throughput to identity. They preserve continuity not only of energy but of phenotypic possibility, making them especially important in differentiation, cancer, and adaptive remodeling [5][7].
4.5 · α-ketoglutarate, succinate, and fumarate — fate and epigenetic continuity Few examples better illustrate the depth of continuity logic than α-ketoglutarate, succinate, and fumarate. These metabolites influence dioxygenase-dependent chromatin and DNA-modifying systems, thereby shaping epigenetic configuration and cell-fate option space. In practical terms, they are loops between metabolic flux and developmental or pathological trajectory. They show that rewiring is not only about changing reaction routes; it can also alter the rules by which future rewiring becomes biologically permissible [5][7].
5 · Layered feedback, community entanglement, and network plasticity
Continuity logic scales upward. It does not stop at the level of a single intracellular pathway. Layered feedback provides one bridge to larger organizational scales, because it allows fast local correction to coexist with slower strategic remodeling. That duality is crucial for networks that must remain robust under repeated disturbance yet retain the capacity to adapt [2].
At the community scale, continuity can also emerge between organisms. Experimental evolution of interkingdom microbial communities has shown that selection can strengthen pre-existing metabolic interactions and even produce de novo interdependence, including around nitrogen metabolism, through pleiotropy and regulatory trade-offs rather than explicit group-level planning [8]. In Continuity Nodes terms, interspecies metabolite exchange can itself become a loop-bearing node: a cross-boundary governor of community viability.
This is strategically important because it suggests that evolutionary rewiring often follows the path of least discontinuity. New architectures are stabilized when they can be threaded through existing loops of energy, redox, nutrient exchange, stress response, and regulatory memory. Rewiring that ignores continuity tends to be lethal; rewiring that co-opts it can become evolutionary infrastructure.
6 · Implications for artificial-life interfaces and adaptive network design
The Continuity Nodes framework is not confined to biochemistry. It offers a design grammar for any adaptive network, including artificial intelligence, hybrid bio-digital systems, and synthetic ecologies. In each case the problem is similar: how can a network reconfigure under pressure without losing viable coherence? The answer is unlikely to be mere scale. It is more likely to be the placement of loop-bearing control sites that preserve state, redirect flux, and transform local error into global correction.
This is why Continuity Nodes matter for the evolutionary dynamics of networking rewiring. They identify the specific places where adaptation can happen without annihilating the system that is adapting. The stronger the continuity architecture, the more freedom the network has to explore new topologies, metabolic regimes, or interface arrangements.
Read this way, metabolic maps become exemplary rather than merely descriptive. They teach that real intelligence—whether cellular, organismal, or synthetic—is not made only of connections. It is made of loops that keep passage viable while the graph is still becoming something else.
SignalSense conclusion. Continuity Nodes are the cybernetic invariants that make rewiring survivable. In metabolism, they appear wherever flux, sensing, memory, and phenotype are tightly coupled. In evolutionary terms, they are not the refusal of change, but the architecture that allows change to persist.
SignalSense · Continuity Nodes Monograph · 5
Selected references
[1] KEGG PATHWAY Database and related KEGG documentation: manually drawn pathway maps representing molecular interaction, reaction, and relation networks; reference and organism-specific pathways. KEGG, accessed 2026. [2] Hu CY et al. Layered feedback control overcomes performance trade-off in synthetic biomolecular networks. Nature Communications. 2022. [3] Choe D et al. Adaptive laboratory evolution of a genome-reduced Escherichia coli. Nature Communications. 2019. [4] Metallo CM, Vander Heiden MG. Understanding metabolic regulation and its influence on cell physiology. Molecular Cell. 2013. [5] Martínez-Reyes I, Chandel NS. Mitochondrial TCA cycle metabolites control physiology and disease. Nature Communications. 2020. [6] Yamada T, Bork P. Evolution of biomolecular networks: lessons from metabolic and protein interactions. Nature Reviews Molecular Cell Biology. 2009. [7] Li X, Egervari G, Wang Y, Berger SL, Lu Z. Regulation of chromatin and gene expression by metabolic enzymes and metabolites. Nature Reviews Molecular Cell Biology. 2018. [8] Scarinci G et al. Enhanced metabolic entanglement emerges during the evolution of an interkingdom microbial community. Nature Communications. 2024.

SignalSense · Continuity Nodes Monograph · 6
📝 About this HTML version
This HTML document was automatically generated from the PDF. Some formatting, figures, or mathematical notation may not be perfectly preserved. For the authoritative version, please refer to the PDF.